The ruminant feedstock provides the growth substrate for all
the micro-organisms that rely on fermentation and in the ultimate analysis
also provides all the growth substrates on which other species of micro-organism
depend; whether they are autotrophs or heterotrophs.
The feed components of interest in this teaching unit are plant carbohydrate
polymers. The two broad divisions of these compounds are the soluble and
insoluble polymers.
Soluble polymers include the starches which are nutrient stores in plants.
The insoluble polymers include cellulose and the pectins and hemicelluloses
which are structural components of plant material. The insoluble, structural
carbohydrate polymers are called plant fibre. Plant fibre in combination
with the largely indigestible heterogeneous structural material called
lignin, and the glycoprotein, extensin, make a rigid and strong structure
that is insoluble in aqueous media.
Ruminants grazing mixed populations of plant pasture species consume
a diet containing a high proportion of plant fibre.
The nutritional use of plant fibre.
 |
Cats and other carnivores are almost completely
unable to take nutritional advantage of plant fibre. Non-ruminants, and
in particular carnivores, may eat diets with a relatively low plant fibre
content but whose starch or other a-linked glucose
polymer content (such as glycogen) may be high. Glucose polymers containing
a-glycosidic
links can be hydrolysed by digestive enzymes such as amylase synthesised
in the mammalian pancreas and carnivores may obtain a high proportion of
daily glucose needs from this source.
Plant fibre in contrast can not be degraded by hydrolases elaborated
by animal cells and can not be used as a nutrient but plant fibre does
serve a useful function in the carnivore diet; it is a bulking agent important
for stool formation.
This is Natalie's cat Togie. |
Herbivores however can use plant fibre as a nutrient but different species
vary in their ability to do so. Ruminants are remarkable for their
ability to utilise plant fibre very well. Even diets high in really intractable
fibrous feed materials are able to be used relatively well in ruminants.
If you examine faeces of different herbivore species you can glean some
indication of the extent to which plant fibre is utilised as a nutrient
source. Ruminants typically produce faeces which do not contain obvious
structural plant material. Contrast this with horse faceces which even
on good quality hay show obvious evidence of the fibre content of the diet.
The reason is that ruminants have a much more agressive digestive system,
specifically designed to enable them to thrive on coarse feed materials.
As we have just seen the soluble a-linked
glucose polymers like starch can be digested by pancreatic amylase to release
glucose for absorption in the small intestine. The main chemical difference
between soluble and insoluble plant carbohydrate polymers is that the insoluble
carbohydrate contains sugars joined together mostly with
b-glycosidic
ether links. These linkages can not be hydrolysed by enzymes synthesised
in mammalian tissues. They can however be hydrolysed by many bacterial
and fungal hydrolases and ruminants harbour multiple species of these microorganisms,
in symbiotic association, in their greatly expanded anterior gut; the structure
known as the reticulo-rumen.
 |
This Brahman steer is an example of the nutritional
importance of symbiotic micro-organisms in primary production. |
These symbiotic micro-organisms synthesise hydrolase enzymes which they
secrete into the growth medium where they degrade both a
and b -linked dietary carbohydrate polymers.
These hydrolases catalyse the release of monosaccharides from starches
and from cellulose, pectins and hemicellulose which are then available
for use as the growth substrates for many of the rumen micro-organisms.
These substrates are mostly fermented to obtain ATP for growth and the
volatile fatty acids propionate,
acetate
and butyrate,
plus lactate are generated as end-products.
These organic acids are waste products of microbial metabolism but are
absorbed by the host ruminant and used as substrates in the host tissues
for intermediary metabolism.
This use of alpha linked polymer by the micro-organisms precludes glucose
release by amylase digestion in the upper small intestine and this means
that gluconeogenesis occurs constantly in the liver of adult ruminants
to compensate for the lack of glucose taken up from the upper small intestine.
Details of plant fibre chemistry
Cellulose
Cellulose is the most abundant organic compound in the biosphere. It
is an unbranched polymer of a-D glucose joined by 1-4 b-glycosidic
ether links. Digestive enzymes elaborated by mammalian tissues can not
degrade cellulose and it can only be of nutritional benefit to animals
that have gut microorganisms that make cellulases that release the glucose.
The glucose released in this way however is the growth substrate for the
microorganisms which obtain their ATP by fermenting it. The host ruminant
absorbs the end products of fermentation for use as nutrients.
The diagram below shows three glucose monomers linked by 1-4 b-glycosidic
ether linkages in the structure of cellulose. Cellulose fibrils are
made up of many thousands of glucose molecules linked end to end in this
way.
The image below shows how the cellulose fibrils are aligned to make
up sheets which are arranged in overlapping structures to make up plant
cell walls.

The image below gives a closer view

Other components of plant fibre
Hemicelluloses are short, linear polymers of hexose and pentose sugars
on a 1-4 b-polyxylose core. Examples are the
xylans and glucomannans. Like cellulose these polymers can only be of nutritional
benefit to the host after microbial fermentation of the component monosaccharides
and absorption of the end products.
Xylan
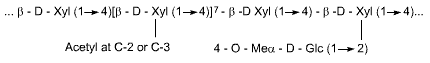
Glucomannan
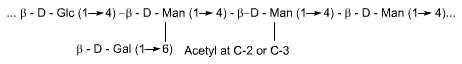
Pectins are 1-4 b-galacturonic acid
polymers with short branches of mixed hexose and pentose sugars. They are
most abundant in soft plant tissues and their sugar content is released
by depolymerising pecitinases, both hydrolases and lyases, produced by
a number of species of organisms.
Lignin is a complex, highly cross-linked heterogeneous polymer
of coumaryl alcohol and its methoxy derivates sinapyl and coniferyl alcohols.
Plants synthesise these compounds from phenylalanine. Lignin exists in
close association with the insoluble plant fibre materials discussed above
and adds to their support functions. Lignin is highly resistant to
chemical and enzymic hydrolysis and for this reason it is not able to be
digested and is of no nutritional value. More important however is
the fact that because it is closely interwoven with the carbohydrate polymer
plant fibre structures it significantly reduces fibre digestibility when
it is present in high abundance. This is one of the reasons for the
low nutritional value of straw.
Extensin is a glycoprotein rich in hydroxyproline residues that
are attachment sites for carbohydrate side chains. It is thought to form
a network complementary to cellulose with cellulose microfibrils penetrating
the extensin matrix.